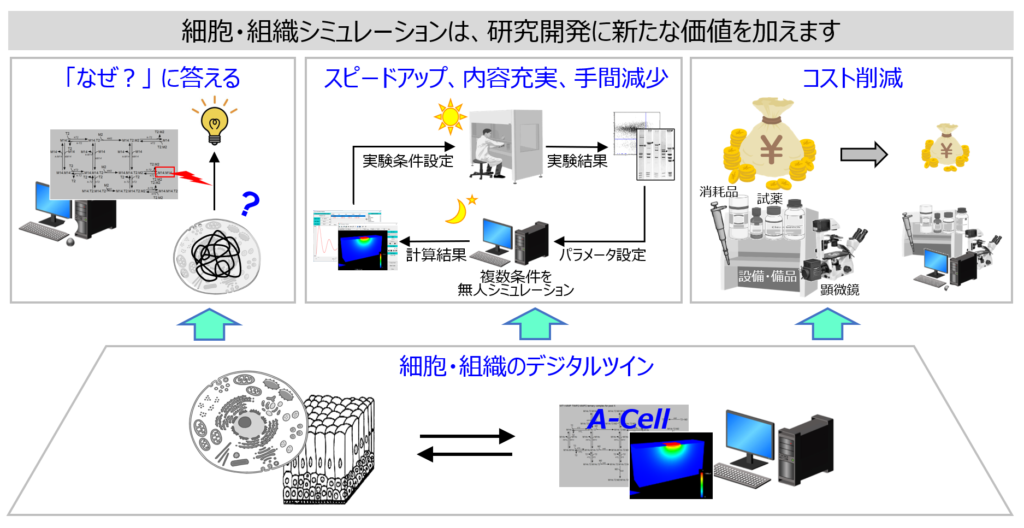
細胞シミュレーションは「なぜ?」に答え、研究開発内容を深め、効率化もします。
細胞シミュレーションには、すべてのパラメータを変更できるという実験に無い特徴があります。この簡単で単純な特徴が細胞の「なぜ?」に答え、研究開発のスピードアップ、内容充実、手間減少、コスト削減を可能にします。例えば実験では標的タンパク質の活性制御にinhibitor等を使いますが、手間と時間と費用がかかります。一方、細胞シミュレーションでは濃度を変更するだけなので、研究開発を効率化できます。
しかしそれだけではありません。複数タンパク質の同時制御など、実験では困難なこと・不可能なことを実現して予測を可能にします。これが新たな価値を生みます。
弊社は、日本はもちろん、世界的にもその最初期(1980年代)から細胞シミュレーションによる研究を開始し、直感的な操作の細胞シミュレーションソフトウエアA-Cellを開発し、ノウハウを蓄積してきました。この経験を活かして、細胞シミュレーションの威力を最大限に発揮させるA-Cellと豊富なノウハウをご提供します。
NEWS
2024年4月1日 「実細胞のデジタルツインをコンピュータの中につくります」をプレス発表しました。これはPR TIMES社様のプロジェクトApril Dreamに賛同して、発信したものです。上記青字のクリックにより、内容をご覧いただけます。
2023年10月24日 「細胞シミュレーションは研究開発を効率化する」をテーマにウェビナーを開催しました。青字のクリックにより、YouTubeでご覧いただけます。
2023年9月25日 10月24日、11:00~11:45開催のウェビナー「細胞シミュレーションの有効性とA-Cell」のメディア発表を行いました。視聴ご希望の方は、右上の「ウェビナー案内・申込」ボタンからお申込みください。
2023年9月11日 10月24日、11:00~11:45開催のウェビナーのお申込みができるようになりました。右上の「ウェビナー案内・申込」ボタンからお申込みください。詳しい内容もご覧いただけます。
2023年9月1日 10月24日、11:00~11:45にウェビナーを開催します。 なぜ細胞シミュレーションは研究開発をスピードアップし、コストを削減し、実験ではできないことができるのかをテーマとして、弊社のお客様からも直接お話いただきます。参加無料です。詳細が決まり次第、本ホームページ上でお知らせいたします。
2023年7月1日 新サービス「お試しA-Cell・コンサルティング」を始めました。左記リンク、右側のボタン、あるいは「コンサルティング→お試しA-Cell・コンサルティング」から概要をご覧になれます。
2021年7月10日 メインテナンス終了のお知らせ: ご迷惑をおかけしておりましたが、メインテナンスが終了しました。このメインテナンスにより、Google Chromeにおけるカード決済の不具合を修正しました。一部のお客様にはご迷惑をおかけして、大変申し訳ありませんでした。
2021年7月10日 重要なお知らせ: 7月10日 14:00~24:00までメインテナンスを行います。復旧までお待ちいただくようお願い致します。
2021年6月30日 重要なお知らせ: 現在カード決済による製品販売を停止しております。お客様には大変ご迷惑をおかけしますが、銀行振込によるご購入をお願い申し上げます。現状復帰が完了したらホームページ上でお知らせ申し上げます。ご不便をおかけすることを重ねてお詫び申し上げます。
2019年8月12日 代表取締役の市川が、長野県須坂市の浄運寺にて、「ウジが湧くの真理」と題して、生物学がどのように真理に迫って来たかについて講演しました。一般の方々120名ほどがお集まりになり、とても熱心に聞いて下さいました。
2019年7月11日 A-Cellで生成したシミュレーションプログラムを、3クリックでコンパイルと実行ができるA-CellRunSimをリリースしました。登録不要、しかも無償です。右のバナーに詳しい説明ととダウンロードボタンがあります。A-CellRunSimで快適なシミュレーション環境をお楽しみ下さい。
2019年3月5日 16:00~17:00の1時間、理化学研究所において、理研セミナー”True Cell Simulation”を行いました。細胞シミュレーションの現状と課題、そして未来に向けた展望について講演しました。
2018年3月23日 小冊子「10ページで知るin silico実験:Simulated FRAP」をリリースしました。どなたでも無償でダウンロードしていただけます。FRAPの実験とシミュレーションは同じ物理的理論に裏付けられているため、シミュレーションはin silico実験であり、wetの実験と比較することが可能です。4D細胞シミュレーション ができるA-Cellの特徴を生かして、FRAPのシミュレーションを行うことにより、wet実験のパラメータを事前に検討したり、実験結果の妥当性を評価したり、実験の精度を上げたりすることが可能になります。是非ご一読いただき、A-CellによるSimulated FRAPをご検討下さい。なお動画もYouTubeで覧になれます。
2018年1月22日 実験医学(羊土社)の連載「予言するシミュレーション」もいよいよ最終回(第7回)となりました。最終回は「予言を生むために:まとめと薬剤ターゲット同定への活用例」です。これまでは事例に応じて予言のコツを述べてきましたが、最終回では、より普遍的な視点からまとめました。また、より切実な問題に予言を適用した事例として、薬剤ターゲットの同定を試みた事例を紹介しています。A-Cellモデルは無償でダウンロードしていただけます。
2017年12月22日 実験医学(羊土社)の連載「予言するシミュレーション」の第6回として、「粒子シミュレーションで細胞のストレス応答を予言する」が出版されました。第5回までは、タンパク質の量を表すのに濃度を用いていましたが、細胞シミュレーションではこれが不適切な場合があります。細胞へのストレス印加によってストレス顆粒が細胞質に分散して形成されますが、その数は数十個程度です。そこで粒子シミュレーションを行い、それによってストレス顆粒の形成ダイナミクスについて予言を行い、ウェット実験で検証した事例を紹介しています。A-Cellモデルは無償でダウンロードしていただけます。粒子シミュレーションは現在のA-Cell範囲を超えていますが、お使いになりたい場合はこちらからお問い合わせください。
2017年11月28日 実験医学(羊土社)の連載「予言するシミュレーション」の第5回として、「細胞応答の柔軟性をパスウェイシミュレーションで予言する:MAPK経路の例」が出版されました。同じパスウェイでも細胞応答が柔軟に制御できることをシミュレーションで予言して、それを実験で検証したBhallaらの事例を紹介しています。論文データからできる限り再現したA-Cellモデルを無償でダウンロードしていただけます。是非とも製品版(Basic EditionあるいはStandard Edition)をご購入いただき、ご自身の研究にご活用いただくようご案内申し上げます。
2017年10月19日 実験医学(羊土社)の連載「予言するシミュレーション」の第4回として、「転写因子NF-κBの核膜輸送による制御を4Dシミュレーションで予言する」が出版されました。A-Cellモデルは第3回と同じですが、今回は核膜輸送による核内NF-κBの振動制御についての予言を紹介しています。NF-κBの核膜輸送自身は振動パターンに影響を与えず、mRNA(IκB)の核外輸送と新規合成を含むIκBの核内輸送だけが影響を与えるという、予想外の予言です。第3回と同様に4Dシミュレーションです。連載で使用したA-Cellモデルは無償でダウンロードしていただけます。是非とも製品版(Standard Edition)をご購入いただき、ご自身の研究にご活用いただくようご案内申し上げます。
2017年9月22日 実験医学(羊土社)の連載「予言するシミュレーション」の第3回として、「転写因子NF-κBのオルガネラによる制御を4Dシミュレーションで予言する」が出版されました。細胞外の刺激を核に伝達するためには、活性化したタンパク質が3D細胞質内を伝わって核に到達しなければなりません。逆に、転写因子による転写制御の効果が細胞質に現れるためには、mRNAや新生タンパク質が核やその近傍から3D細胞質内を移動しなければなりません。今回はこのような状況をNF-κBでシミュレーションし、得られた予言についてご紹介しています。是非ご一読いただくようご案内申し上げます。連載で使用したA-Cellモデルは無償でダウンロードしていただけます。
2017年8月21日 弊社が執筆を担当している実験医学(羊土社)の連載「予言するシミュレーション」の第2回として、「神経細胞間の信号伝達効率:どのように制御されるのか?」が出版されました。今回はパスウェイモデルによる予言です。是非ご一読いただくようご案内申し上げます。連載で使用したA-Cellモデルは無償でダウンロードしていただけます。
2017年7月12日 弊社代表取締役・市川一寿が執筆する実験医学(羊土社)の新しい連載「予言するシミュレーション」が始まりました。シミュレーションの最も大事な役割の一つは予言です。なぜならシミュレーションによる予言と実験を組み合わせることによって、素早く仮説・検証サイクルを回すことができるからです。また予言はシミュレーションが最も得意とすることでもあります。連載第1回は「4Dシミュレーションでがん浸潤阻止の新しい方法を予言する」です。是非ご一読いただくようご案内申し上げます。なお連載で使用したA-Cellモデルは無償でダウンロードしていただけます。
2017年5月8日 Windows 10でA-Cellが起動できない例が報告されています(フリーエディションを含む)。これを回避するパッチをリリースしました。これに伴い、すべてのA-Cellエディションの変更を行いました。
2017年3月10日 A-Cellには分子細胞生物学の研究での利用、薬剤開発の研究での利用、そして大学教育での利用の3つの利用法があります。このたび大学教育での利用について、17例をご紹介した小冊子「大学教育でのA-Cell利用例」をリリースしました。左記リンクから、あるいはA-Cellページからご覧いただけます。これがすべてではないことは言うまでもなく、今後利用例を追加して行く予定です。
2017年3月7日 「NF-κBの4D細胞シミュレーション」と題する記事が、実験医学増刊号(2017、35巻、p.166、羊土社刊)に掲載されました。
2016年11月27日 細胞シミュレーションの例として、タンパク質のダイナミクスを測定するFRAP(蛍光退色後回復測定)をA-Cellでシミュレーションした例を、YouTubeでご覧いただけます(”Simulated FRAP by A-Cell“、17分程度)。これをご覧になれば、4D細胞シミュレーションのモデルの構築から、結果を表示するまでの様子も同時に知っていただけます。
2016年10月21日 高性能コンピューター(HPC)の開発販売を行っておられるHPCシステムズ株式会社様による弊社のインタビュー記事が掲載されました。4D細胞シミュレーションをスピーディーに行うためにはHPCが不可欠で、これまでの研究にも多用してまいりました。このインタビューでは4D細胞シミュレーションについて分かり易くまとめられていますので、是非ご覧ください。
2016年8月31日 小冊子「10ページで理解する4D細胞シミュレーション」をリリースしました。どなたでも無償でダウンロードしていただけます。これは「10ページで理解するパスウェイシミュレーション」の姉妹版で、パスウェイシミュレーションを細胞内の広がり(空間)に拡張してシミュレーションを行うための基礎とメカニズム、及びその必要性について解説しました。4D細胞シミュレーションは、今後重要性を増すと考えられます。是非ご一読下さい。
2016年7月6日 「細胞シミュレーションサービス」を開始しました。ご提供いただく実験結果に基づいて弊社がシミュレーションを行い、実験と比較してシミュレーション結果とモデルファイルなどをお返しします。これにより実験の確認を行ったり、問題の事前推定などを行うことができます。上記のトップメニューから細胞シミュレーションサービスを選択して詳細をご覧いただけます。
2016年6月17日 小冊子「A-Cellを使ったパスウェイシミュレーション ERKシグナル伝達」をリリースしました。これは10ページシリーズに加えて、無償でダウンロードしていただける新たなシリーズです。すでに発表された論文のシミュレーションをA-Cellで再現する方法を解説しています。モデルファイルやサンプルシミュレーション結果もダウロードしていただけます。今回はERKシグナル伝達ですが、他のパスウェイについても今後発刊予定です。
2016年6月17日 論文発表されている細胞シミュレーションをA-Cellで再構築するサービスを開始しました。トップメニュのA-Cell>A-Cellによる論文シミュレーションの再構築サービス、からご依頼・お問い合わせをお願い致します。
2016年4月20日 A-Cell講習会(第3回)を5月30日(月)に東京で開催します。今回は個別相談の時間を設けましたので、各研究固有のご相談をしていただけます。なお、A-Cellご購入者は無料で受講いただけます。
2016年4月7日 A-Cellご購入の際に、クレジットカードに加えて、銀行振込でお支払いただけるようになりました。これは見積→発注→納品→請求の流れに沿って、購買部門ご担当者経由で購入いただける方法です。科研費などの公費にも対応致します。
2016年3月25日 「A-Cell入門」の新しいバージョンをリリースしました。
2016年3月17日 3月15, 16日にグランドハイアット東京で開催されたバイオアジア2016において、A-Cellと細胞シミュレーション・コンサルティングのプリゼンテ―ションを行いました。皆様のご来場ありがとうございました。
2016年1月14日 小冊子「10ページで知る細胞シミュレーションの活用法」を発行しました。自分の研究にシミュレーションがどのように活用できるかのヒントを得ていただくため、8つの活用例を紹介しています。無償ダウンロードができますのでぜひご一読下さい。
2015年12月28日 A-Cellのご購入時に、コンビニでのお支払が可能になりました。
2015年11月25日 A-Cellの販売を開始しました。使える機能が異なる3つのエディション(エントリー、ベーシック、スタンダード)をご用意しました。学生さんからプロの研究者まで、幅広くご利用いただけます。A-Cellの素晴らしい機能と使い心地を、是非ともご活用下さい。
2015年11月25日 A-Cellの販売開始を記念して、A-Cell無料講習会を開催致します。 A-Cellの基本的な使用法を解説しますので、ご購入済の方はもちろん、ご検討中の方にもきっとお役に立つ内容です。奮ってのご参加をお待ち申し上げます。
2015年11月19日 「A-Cell操作法」がダウンロードできるようになりました。A-Cellをご購入された方がダウンロードできます。A-Cellはまもなくご購入いただける予定です。
2015年11月12日 小冊子「10ページで理解するパスウェイシミュレーション」を発行しました。パスウェイシミュレーションに興味を持っている方や、細胞の実験を専門にしつつ概要を知っておきたい方などを対象に書きました。寝っころがって読めることを心掛け、無償ダウンロードができますのでぜひご一読下さい。
2015年10月23日 11月16日(月)に第2回「A-Cell初級講習会」を開催します。今回は、シミュレーションによって何が得られてどのような進展が期待できるのか、実験をシミュレーションに結び付けるにはどうすれば良いのか、また実験の中にどう位置づけたら良いのかを中心に解説します。詳しくは右のバナーからご覧ください。
2015年10月20日 Springerから書籍が出版されました。1) Ohshima, D., et al., “Potential Roles of Spatial Parameters in the Regulation of NF-κB Oscillations, as Revealed by Computer Simulations”, pp63、2) Ohshima, D., et al., “Stochastic Simulation of Stress Granules”, pp.77. いずれも書籍”Protein Modifications in Pathogenic Dysregulation of Signaling”, eds. Inoue, J-I. & Takekawa, M., Springer, Tokyo, 2015, ISBN 978-4-431-55560-5, ISBN 978-4-431-55561-2 (eBook)中の2章です。
2015年10月14~16日 10月14日(水)~16日(金)の3日間、パシフィコ横浜で開催されたBioJapan2015に出展しました。ブースB103での展示と、10月15日(木)10:20-10:50、C会場におきまして「細胞シミュレーションを武器にした新たな創薬戦略」と題してプリゼンテーションを行いました。プリゼンテーションへのご参加、またブースへお立ち寄りいただき、まことにありがとうございました。
2015年10月4日 英語版ホームページを立ち上げました(http://engl.tc-simulations.com/)。ご活用いただくようご案内申し上げます。
2015年8月26日 8月24日にA-Cell初級講習会を開催しました。論文のConclusionなどに掲載されるシグナル伝達のポンチ絵を基にしてモデルを創り、シ ミュレーションの実行とグラフ表示ができるようになることが講習会の目標でしたが、参加者全員が目標を達成されました。ハードル高く感じていた細胞シミュ レーションですが、A-Cellを使えば簡単に実行できることを実感いただけたと思います。次回開催を予定していますので、この機会を逃した方々のふるってのご参加をお待ちしています。(当日の開催概要はこちら)
2015年8月7日 8月6日に製薬企業をはじめ様々な企業や大学からの研究者に加えて各界でご活躍の方々にも参加いただき、シンポジウム「実験-数理細胞シミュレーション共同研究の新たな局面:創薬展開への可能性」を開催しました。実験側の先生と数理・シミュレーション側の先生が組になって同じ研究テーマを発表する、というこれまでにないスタイルを企画しましたが(全5テーマ)、これによって数理・シミュレーションになじみの薄い方々にも実験と共同して数理・シミュレーション研究を進める方法を知っていただきました。最後のパネルディスカッションでは数理・シミュレーションの意義に迫る本質的質問がパネリストや会場から出され、真剣で緊張感あふれるやり取になりました。第2回目の開催を望む声が聞かれる中、盛況のうちに終了しました。(当日のプログラムはこちら)
2015年6月29日 論文“Spatio-temporal dynamics and mechanisms of stress granule assembly”がPLoS Computational Biologyに掲載されました。細胞が低酸素状態になったりヒ素などの薬物暴露があったりするとストレス顆粒(stress granule)が形成され、細胞の運命決定に重要な働きをします。この論文では、ストレス顆粒の細胞内ダイナミクスを初めて明らかにしました。論文で使った確率的シミュレーションのA-Cellへの実装を現在進めています。(この論文で使ったA-Cellモデルと論文中で使った確率的シミュレーション プログラムはこのダウンロードボタンからどうぞ。)[wpdm_file id=18]
2015年6月18日 論文”Synergistic effect of blocking cancer cell invasion revealed by computer simulations”がJournal of Mathematical Biosciences and Engineeringにアクセプトされました。ECM分解の抑制法とMMP-2の役割について議論しました。(この論文で使ったA-Cellモデルはこのダウンロードボタンからどうぞ。)[wpdm_file id=15]
2015年6月4日 論文“Regulation of NF-κB oscillation by nuclear transport: Mechanisms determining the persistency and frequency of oscillation”がPLoS ONEに掲載されました。この論文では転写因子NF-κBの核内振動が核膜輸送でどのように制御されるか、また制御のメカニズムは何かを初めて明らかにしました。(この論文で使ったA-Cellモデルはこのダウンロードボタンからどうぞ。)[wpdm_file id=16]
2015年4月21日 論文“Regulation of NF-κB oscillation by nuclear transport: Mechanisms determining the persistency and frequency of oscillation”がPLoS ONEにアクセプトされました。
2015年4月8日 論文“Spatio-temporal dynamics and mechanisms of stress granule assembly”がPLoS Computational Biologyにアクセプトされました。